La desmielinización
es un proceso patológico en el cual se daña la
capa de mielina de las fibras nerviosas. La pérdida de
las vainas de mielina en los axones de las neuronas es el distintivo
de las llamadas enfermedades desmielinizantes; esta destrucción
puede implicar el mal funcionamiento de órganos o músculos.
Cuando la mielina es destruida, la conducción de las
señales a lo largo de los nervios se ve seriamente afectada,
en consecuencia, los nervios se marchitan con el tiempo. La
desmielinización comporta asimismo una pérdida
de velocidad de conducción del impulso y de respuesta,
pérdida de las propiedades de conducción, consumo
energético poco eficiente y desorganización del
sistema nervioso.
Causas
de la desmielinización
La desmielinización
puede ser causada por una gran variedad de desórdenes
infecciosos, genéticos y metabólicos. A pesar
de tener causas diferentes, la desmielinización se presenta
de forma similar en su desarrollo patológico, dificultando
el diagnóstico médico oportuno de la enfermedad.
Infecciosas
La pérdida de mielina puede estar asociada a infecciones
debido a respuestas del sistema inmunitario, que las considera
como sustancias ajenas al organismo. Un ejemplo es el caso de
la Encefalomielitis Aguda Diseminada (EAD), que normalmente
comienza entre una y cuatro semanas después de una vacunación
o por sangre o de una infección sistemática, o
el caso de la Encefalomielitis aguda hemorrágica (de
W. Hurst), que sucede después de una infección
respiratoria.
Genéticas
La causa puede ser genética si se ven alterados los genes
que codifican proteínas de la mielina impidiendo su normal
producción, como en el caso de las leucodistrofias o
en la enfermedad de Pelizaeus-Merzbacher.
Metabólicas
Otra posibilidad es la afectación de moléculas
implicadas en el metabolismo tales como las enzimas. Un ejemplo
es la Leucodistrofia Metacromática en la que pueden verse
afectadas hasta tres enzimas (la Arilsulfatasa A, multiples
sulfatasas y el activador sulfátido o saponina).
Composición de la mielina
La mielina es
particularmente rica en fosfolípidos. La organización
molecular de la mielina ha sido estudiada mediante microscopía
electrónica. Estos estudios revelan que las capas alternantes
de lípidos mixtos y proteínas corresponden a capas
densas que rodean el axón formando un espiral. Estas
capas están formadas por unas células especiales,
llamadascélulas gliales. Dependiendo de si la mielina
es sintetizada en los axones del Sistema Nervioso Periférico
o del Sistema Nervioso Central, la célula glial que lo
forme será una célula de Schwann o un oligodendrocito respectivamen-
te.
Las membranas de mielina son ricas en esfingolípidos
y tienen gran cantidad de glucoesfingolípidos.
Aspectos biológicos
Las células mielinizadas tienen una mayor velocidad de
transmisión y una conducción más eficiente
respecto a las no mielinizadas. Por lo tanto, los axones pueden
ser más finos en los organismos que presentan células
mielinizadas. La fineza de los axones permite que el sistema
nervioso pueda organizarse adecuadamente ocupando el menor espacio
posible. En consecuencia, se estima que gracias a la mielinización
el cerebro humano es diez veces más pequeño de
lo que sería sin mielina en sus fibras nerviosas y que
además el gasto metabólico que implica su funcionamiento
es también diez veces menor.
Aspectos bioquímicos
Lípidos de la mielina
La mielina es una estructura fuertemente empaquetada debido
a las interacciones hidrófobas entre los lípidos
y las proteínas.
Los cerebrósidos constituyen aproximadamente el 16% de
los lípidos totales de la mielina, siendo el galactosilcerebrósido
el más abundante. Éste está compuesto por
un único azúcar unido al grupo hidroxilo de la
esfingosina. El alto contenido de colesterol de la membrana
también contribuye al fuerte empaquetamiento. Los ácidos
grasos sintetizados por el cerebro no tienen carga en sus cadenas
laterales, lo que hace posible que la mielina sea muy compacta.
La esfingomielina, por su parte, se encuentra en una proporción
baja.
Proteínas estructurales de la mielina
Las capas de mielina se mantienen unidas por interacciones entre
proteínas con lípidos y entre proteínas
entre sí. Cualquier interrupción de estas interacciones
puede conducir a la desmielinización de la membrana.
Las proteínas principales son diferentes según
si hablamos del Sistema Nervioso Central o del Sistema Nervioso
Periférico:
Las proteínas principales del Sistema nervioso Central
son dos. Estas constituyen entre el 60% y el 80% de todas las
proteínas: una es la proteína proteolípido
(PLP), y la otra es la proteína básica de la mielina
(MBP). La PLP es hidrófoba y relativamente resistente
a la proteólisis. Se cree que la PLP tiene la función
de promover la formación y estabilización de la
estructura de múltiples capas de la mielina. A diferencia
de las PLP, las MBP (son un conjunto de proteínas) se
pueden extraer fácilmente de la membrana y son solubles
en disolución acuosa. Estas se encuentran en la cara
citoplasmática de las membranas de mielina.
En el Sistema nervioso periférico la principal proteína
de la mielina es el Po. Esta es una glucoproteína que
representa más del 50% del contenido proteico de la mielina
del SNP. El Po parece tener la función de mantener la
estructura de la mielina.
Aspectos biofísicos
La importancia física de la mielina radica en que funciona
como aislante eléctrico. Esta característica permite
ahorrar energía a la célula ya que la excitación
activa solo se produce en los nódulos de Ranvier; y también
permite aumentar la velocidad de transmisión de la señal,
ya que el potencial de acción se propaga saltando de
un nódulo de Ranvier a otro (conducción saltatoria).
Transmisión de un estímulo
a través de una neurona
Cuando una célula no excitable recibe una despolarización,
esta se atenúa al cabo de un tiempo. En cambio, si esta
despolarización se da en una célula excitable
como una neurona, no se produce atenuación, sino que
se propaga gracias a un pulso de potencial de membrana que se
dispara. A pesar de esto, si el estímulo no supera un
determinado umbral, no se disparará el pulso, y cuando
lo supere, se disparará. Todos los estímulos que
superen el umbral generan un pulso de la misma intensidad. Así
pues, distinguimos estímulos de más o menos intensidad
a partir de la frecuencia de los impulsos nerviosos (un estímulo
más intenso generará impulsos nerviosos con más
frecuencia que otro que no lo sea tanto).
La diferencia de potencial que se mantiene gracias a la actividad
de las bombas iónicas entre la cara interna y externa
de la membrana plasmática es la base para entender como
se propagan los impulsos nerviosos. La cara interna se encuentra
cargada negativamente, pero cuando llega una despolarización
se produce una inversión de la polaridad de la membrana,
y la parte interna se carga positivamente.
Esta despolarización es posible gracias a los canales
iónicos operados por voltaje. Primeramente actúa
el canal sódico: con la llegada de la despolarización,
una proteína con una región hélice a cargada
positivamente que obturaba el canal se ve atraída por
la cara externa de la membrana, ahora cargada negativamente
a causa de la inversión de la polaridad. El desplazamiento
de la proteína permite dejar libre el canal sódico,
de forma que se produce un influjo de iones de sodio hacia el
interior de la célula. El aumento de permeabilidad del
sodio implica que la membrana pase a tener un potencial muy
próximo al potencial de Nernst del sodio, de valores
positivos.
El canal solo está abierto durante un periodo de tiempo
muy corto (del orden de los milisegundos), y pasado este tiempo
ya no deja pasar más iones. En este momento el canal
permanecerá inactivo durante otro periodo breve, el periodo
refractario, durante el cual el canal no se podrá abrir
ni cerrar. De esta forma se evita que el impulso retroceda.
El aumento de permeabilidad del potasio a través de la
membrana se da justo cuando decae la permeabilidad del sodio.
Este aumento provoca que la membrana adopte un potencial muy
cercano al potencial de Nernst del potasio, de -90mV, lo cual
justifica la hiperpolarización de la membrana. Finalmente
la permeabilidad del potasio también decae y la membrana
de repolariza.
Beneficios de
la mielina
La constante de longitud es la distancia a la cual se ha producido
una atenuación del 63% de la diferencia de potencial.
Ausencia
de mielina
El problema que surge es una disminución de la velocidad
de propagación de la señal.
Otro problema que surge al producirse una desmielinización
es un aumento del uso de energía. Esto se debe a que
la mielina, gracias a su capacidad aislante, bloquea los canales
iónicos regulados por voltaje e impide que funcionen
y que gasten energía. Si la mielina no los recubre, estos
se activarán y consumirán parte de la energía
que produzca la célula.
Estrategias para compensar la falta de
mielina
Algunos animales, como los calamares, poseen unos axones desmielienizados.
Para mantener una constante de longitud alta y evitar la disipación
del impulso nervioso disponen de unos axones muy gruesos.
Enfermedades desmielinizantes
La composición de la mielina es diferente para el sistema
nervioso central y para el periférico. Así pues,
aunque existen procesos inflamatorios o errores congénitos
(de nacimiento) que pueden afectar a la mielina del SNC y del
SNP, también hay enfermedades que solo afectan a una
de estas.
Síntomas generales de la desmielinización
 Los criterios
patológicos más aceptados respecto a
enfermedades desmielinizantes son:
Los criterios
patológicos más aceptados respecto a
enfermedades desmielinizantes son:
• Destrucción de las capas de mielina de las fibras
nerviosas, con indemnidad relativa de otros elementos del tejido
nervioso.
• Infiltración por parte de células de inflamación
en una distribución alrededor de los vasos y
particular- mente al lado de venas.
• Distribución predominante de las lesiones en
la sustancia blanca.
Algunas enfermedades
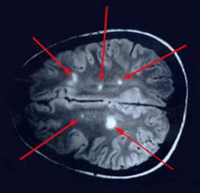
Esclerosis múltiple
Es una enfermedad crónica del SNC que consiste en la
aparición continua de focos de inflamación autolimitados
en zonas de la sustancia blanca (parte del SNC cubierta de mielina)
que pueden dejar como secuela la desmielinización definitiva
de esa zona.
Esta enfermedad se produce cuando el sistema inmune de una persona
ataca sus propias fibras nerviosas mielinizadas, destruyendo
sus propiedades de conducción. Si los nervios dañados
inervan músculos, el movimiento se verá perjudicado.
Leucodistrofia
Son un conjunto de enfermedades genéticas del metabolismo
de la mielina (también se les conoce como enfermedades
desmielinizantes).
Enfermedad de Pelizaeus-Merzbacher Esta enfermedad se debe a
mutaciones en el gen de la proteína proteolipídica
(PLP) en el cromosoma X. El gen afectado es el encargado de
codificar dos proteínas de la mielina del SNC: la proteína
proteolipídica y su isoforma DM20.

